 図1
図1原核生物から真核生物へ
現在地球上にすむ生物は、すべて30億年以上前に生まれたただ1つの原始細胞に由来していると考えられている。この細胞は競争相手よりも繁殖力に優れ、細胞分裂や進化の過程で優位にたち、ついには地球を緑でおおい、大気の組成を変え、この星を知的生物のすみかにまでした。地球上におけるすべての生物間の著しい類似を説明するためにはこの考え方以外にない。進化の道程にはたいへん重要な出来事が約15億年前に起こったと考えられている。つまり内部構造が比較的単純で小さな細胞―いわゆる原核細胞
(procaryotic cell)で、種々の細菌を含む―から、高等な動植物にみられるような大きく複雑な真核細胞が栄えるように変わったのである。
原核細胞は構造的には単純だが生化学的には多様である。
細菌
(bacterium)は自然界に存在する最も単純な生物である。細胞壁をもっている場合が多く、細胞膜があり、DNA、RNA、タンパク質などを含んだ細胞質を包み込んでいる。電子顕微鏡により細胞内部がマトリックスかできているのが分かる。細菌は迅速な分裂能力により環境変化にすばやく適応できる。そして自然界においてはあらゆる生態学的場所(ニッチ)にすんでおり、環境に対応して多様な生化学的性質をもつ。
細菌にはかなり性質の違う
2群、真正細菌と古細菌がある。前者は土や水、生体内など普通の場所にすみ、後者は沼地、深海、塩水、酸性温泉などの悪環境にすんでいる。細菌は簡単な構造をしているが、他のどんな生物よりも地球上に長く生息しており、今でも地球上に最も多く存在する細菌である。
代謝反応の進化
生命が誕生した当初は、比較的簡単な化学反応を行う細胞が周囲の分子を使って生き残り、増殖できたため、代謝反応は必要なかった。しかし進化が進むにつれ限られた天然の資源をめぐる競争が激しくなり、有用な有機物を効率よく作り出す酵素を発達させた生物が選択のうえで有利になった。以来、細胞の持つ酵素の数は次第に増え、現存生物の代謝経路ができてきた。
原始地球の大気中には遊離の酸素はなかったので、最古の代謝経路は嫌気的な解糖であった。解糖はあらゆる生物の細胞で行われており、
ATPを生成する。この単純だが強力な化学反応の活用は、RNAさえも進化する以前の、火山の噴火の硫黄で満ちた原始地球の環境で起こった生命の誕生以前の反応を反映するなごりである。
大気の組成の変化と進化の過程
有機物の合成に必要な物質をめぐる競争が激しくなるにつれ、
CO₂やN₂を直接大気中から利用できる生物が選択上有利になった。だがCO₂やN₂は非常に安定なため、それらを糖など利用できる有機物に転換するには、複雑な化学反応と大量のされる。そして、強い電子供与体がCO₂やN₂の還元に使われる。現存する細菌の光合成機構を比較すると、最初の電子供与体の1つはH₂Sだったと考えられる。その結果蓄積するのは硫黄原子である。後にH₂Oから電子を得るという、困難だがより有利な過程が生まれ、O₂が廃棄物として大量に放出された。酸素を放出する細菌の出現により、大気は分子状の酸素を含まない状態から酸素が全体の21%を占める状態へと変化した。酸素の利用により、生物は摂取した分子をより完全に酸化することができる。それによって得られるATPの量も増える。
共生説
大気中に分子状酸素が蓄積したあと、嫌気性生物は不利な立場に置かれた。絶滅してしまったものもあれば呼吸能力を獲得したものや、嫌気的な生活を続けられるように酸素がほとんど存在しない場所を見つけたものもある。好気性の細胞を捕食するものや、これに寄生するもの、つまり、好気性の細胞と親密な関係を作り、共生を始める一群も現れた。
葉緑体
p684
葉緑体は生化学的な証拠から酸素を生産する光合成細菌が原始的な真核細胞にエンドサイトーシスで取り込まれ、内部に共生したものの子孫と考えられている。
ミトコンドリアも、細菌がエンドサイトーシスで取り込まれたものと一般に考えられている。葉緑体とミトコンドリアがかなり違うのは、先祖となった細菌の違いや、その後の進化における分岐によるのだろう。それにもかかわらず、光で駆動される葉緑体の
ATP合成と、呼吸鎖で駆動されるミトコンドリアのATP合成の基本機構は大変よく似ている。
p684
葉緑体は色素体(plastid)と呼ばれる一群の細胞小器官のうちでもっとも重要なものである。
p685
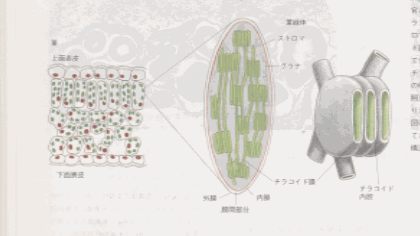
葉緑体は、ミトコンドリアと同じ化学浸透機構を使ってエネルギー変換を行い、同じ原理に基づいて構築されている。外膜は透過性が高いが内膜は低く、そこに特異的な運搬系タンパクが埋め込まれている。内外の膜の間には狭い膜間部分があり、内膜はストロマと呼ばれる広い空間を囲んでいる。ストロマはミトコンドリアのマトリックスに対応し、さまざまな酵素、リボゾーム、
RNA、DNAを含んでいる。葉緑体とミトコンドリアの構造には重要な違いもある。葉緑体内膜はクリステのように折りたたまれておらず、電子伝達系もない。その代わり、偏平な円盤状の袋を積み重ねたチラコイドという第三の膜系があり、その中に電子伝達系をはじめ、光吸収系、
ATP合成酵素がすべて含まれている。チラコイドの内腔どうしは連続しており、チラコイド膜でストロマと隔離されたチラコイド内腔と呼ばれる第三の区画をなしている。
図1
図2
ミトコンドリアと葉緑体の比較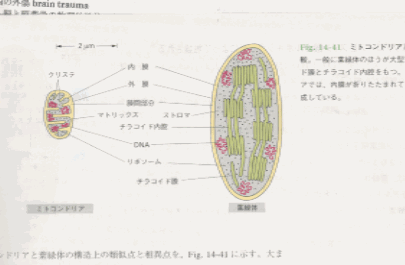 p687
p687
ミトコンドリアと葉緑体の構造上の類似点と相違点を図2に示す。大まかに言えば、葉緑体とはミトコンドリアを非常に大きくし、クリステを切り離してマトリックス中で内部が連続した一連の亜ミトコンドリア顆粒に変えたようなものである。葉緑体の
ATP合成酵素の頭の部分、つまりATPの合成が行われる部分はチラコイド膜からストロマに突出しており、ミトコンドリアのATP合成酵素の頭がクリステの膜からマトリックスに出ているのと同じである。
p691
葉緑体で起こる光合成電子伝達反応に戻ろう。
必要なエネルギーは、クロロフィル分子が吸収した太陽光から得られる。このエネルギー変換の過程では、最初にクロロフィル分子が光量子によって励起され、電子が一個、ある軌道からよりエネルギーレベルの高い軌道へと移る。このような励起分子は不安定でもとの非励起状態に戻ろうとするが、それには三つの方法がある。
光合成反応では、
が利用されている。
p696
葉緑体は、光合成のほかにもいろいろな生合成反応を行う。細胞の必要とするすべての脂肪酸や数種のアミノ酸を、葉緑体のストロマにある酵素が合成する。また葉緑体では、光で活性化された電子のもつ還元力を使って亜硫酸イオン(NO2-)をアンモニア(NH3)に還元する。植物体内ではこのアンモニアが、アミノ酸やヌクレオチドの合成に必要な窒素源となる。このように葉緑体は、植物や藻類で光合成という役割以外にも、代謝上非常に重要な働きをしているのである。
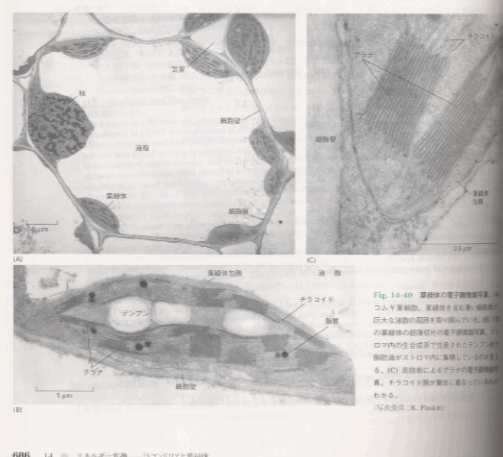
図3
葉緑体の電子顕微鏡
細胞の起源
ミトコンドリアの起源
ほとんどの真核生物で細胞質の体積の相当な部分を占めているミトコンドリアは、複雑な動物が進化し出現するためにも欠かせないものであった。そのミトコンドリアの起源を考えるために、まず背景を溯ってみる。
今からおよそ
35億年以上も前に生じた最初の生きた細胞は、酸素がなく地球科学的な反応でできた有機物が豊富に存在している環境だった。したがってごく初期のATP生産系は現在の発酵とよく似た代謝系だったと考えられる。しかしやがて、これらの代謝活動によって局所的な環境が変化し、さらに新しい生化学的反応経路が進化せざるを得ない状況となった。発酵による排出物が蓄積したために、エネルギーを保存する電子伝達系の進化が促され、嫌気性細菌は非発酵性の有機物をエネルギー源として利用できるようになった。このようにして、細胞内のpHを中性に保ち十分量のエネルギーを確保できるようになったが、細胞の多種多様な分子の前駆体となる糖を作るために、不足しつつある発酵性栄養物質の代わりとなる炭素源を見つける必要が生じた。エネルギー代謝の進化で突破口を開いたのが、太陽光のエネルギーからNADHのような分子を直接作り出せる光化学反応中心の出現であった。これは細胞の進化の初期、おそらく30億年以上前に緑色硫黄細菌の祖先で初めて起こったと考えられている。こうして光合成細菌が無尽蔵に還元力を補給するようになって、細菌の進化における重大な障害が克服された。さらに30億年ほど前に複雑なシアノバクテリアの光合成電子伝達鎖によって大気中に酸素が生じ、好気的代謝によりATPを合成する細菌の出現が可能となった。つまり、光合成の結果、地球上に有機物が蓄積してくると、大腸菌の祖先を含む一部の光合成細菌は光エネルギーだけで生存できる力を失い、完全に呼吸に依存するようになった。このような呼吸依存性の細菌が原始的な真核細胞に入りこんで共生したものがミトコンドリアであり、その出現はほぼ15億年前と考えられている。これが内部共生説(endosymbiont hypothesis)である。これらの生物は、炭水化物や他の還元された有機分子をCO2、H2Oにまで完全に分解し、その時に放出される多量のエネルギーを利用できる。もっと詳しく説明すると、真核生物はミトコンドリアも葉緑体もない嫌気的な生物として出発したあとに、細菌と共生関係を確立し、そのリン酸化系を利用するようになった。ミトコンドリアの進化の発端となったエンドサイトーシスは、大気中にかなりの酸素が蓄積された15億年ほど前、まだ動物と植物が分岐していない時期に起こったと考えられている。その後にエンドサイトーシスにより光合成細菌を取り込んだ植物や藻類が出現した。
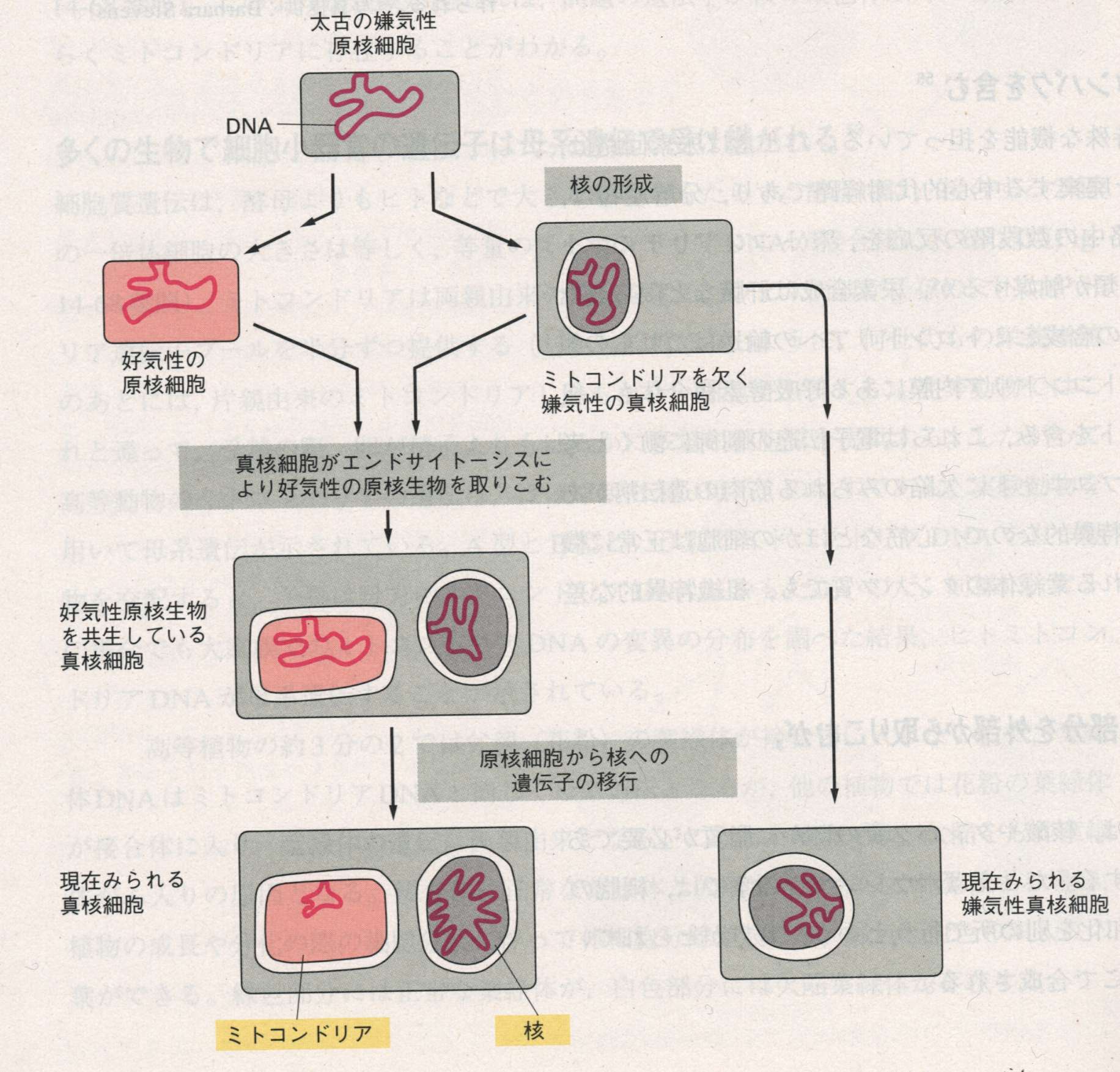
上図はミトコンドリアの起源に関する進化の推定経路である。
酸素呼吸自体は、ほぼ
20億年前に出現したとされ、下図に示すように、酸素呼吸は、緑色細菌、紅色細菌、藍色細菌(シアノバクテリア)系統の光合成細菌で、それぞれ独立に進化したらしい。そのうち光合成能を喪失した好気性の紅色細菌からミトコンドリアが生じたと考えられている。塩基配列の詳しい解析から、ミトコンドリアは、根粒菌、アグロバクテリアやリケッチアに似た細菌から生じたと思われる。これら3種の菌は近縁関係にあり、現存する真核細胞とも、きわめて密接な共生関係を作ることで知られている。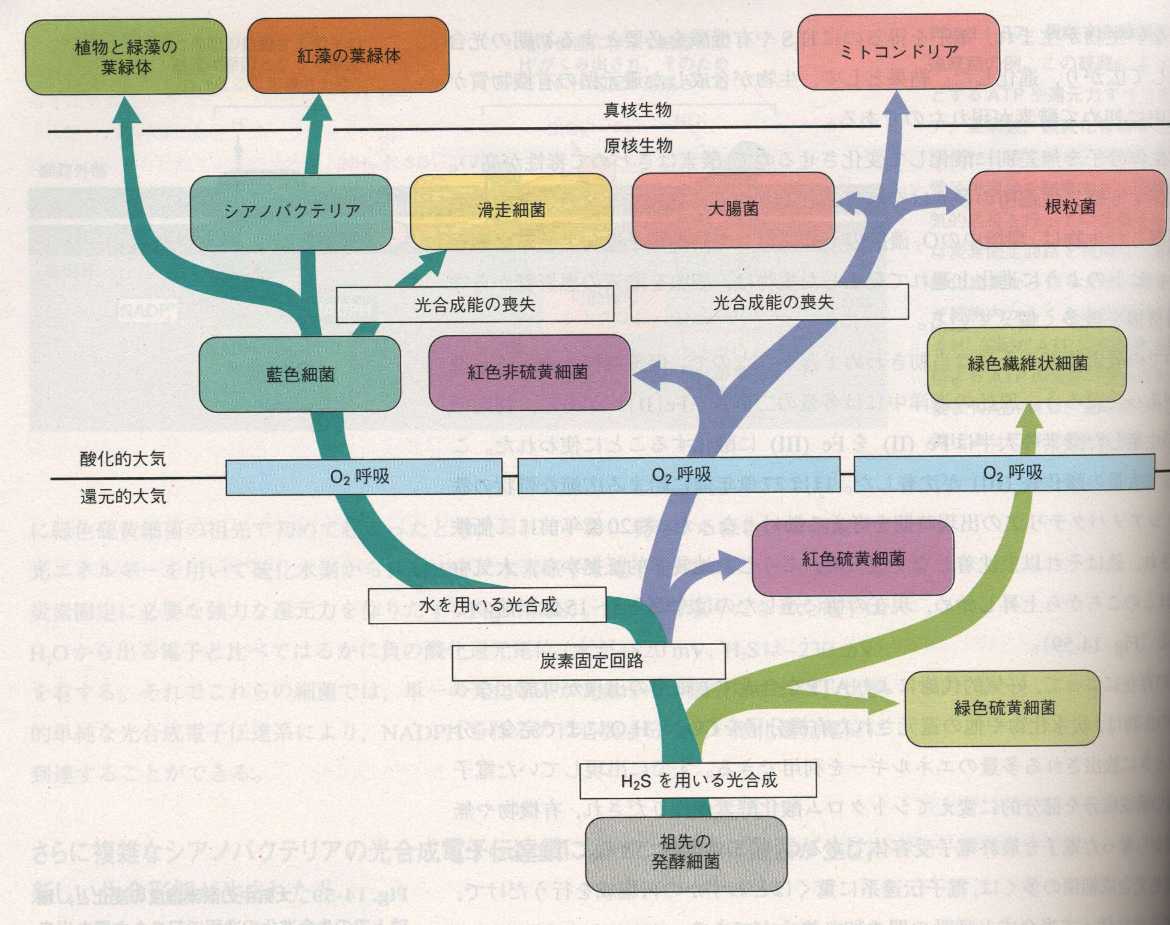
また、細菌、葉緑体、ミトコンドリアには、ミトコンドリアのシトクロムb-c1複合体とよく似た膜結合酵素複合体がある。これらの複合体はいずれも電子伝達体であるキノン(
Qと表している)から電子を受け取り、それぞれの膜を介して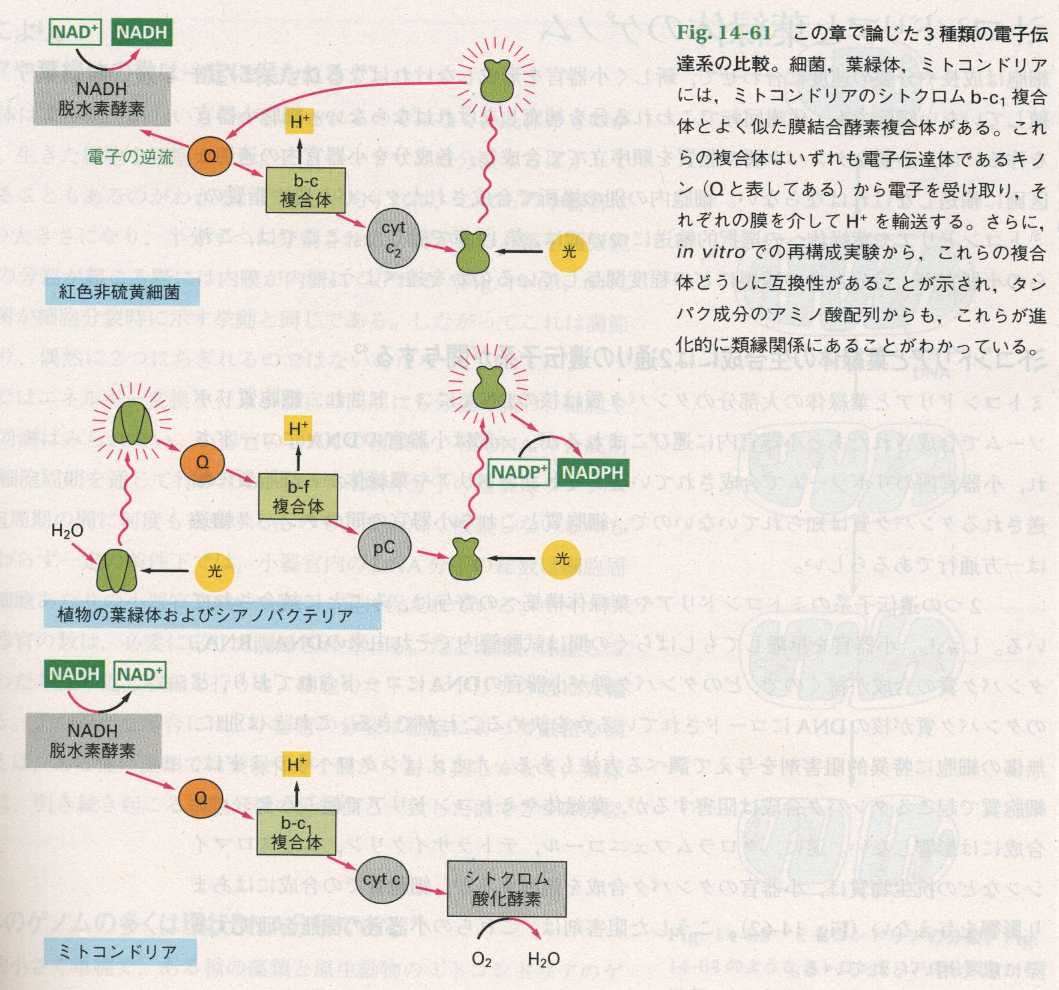
以上のことからミトコンドリアは、特別な紅色光合成細菌が光合成能を失い、呼吸鎖を残したものの子孫であると思われる。しかし、全てのミトコンドリアがただ一度おきた細胞内共生から生じたとは言いきれない。たとえば原生動物のミトコンドリアは原核生物の特徴をはっきりと示すが、植物や動物のものとはかなり異なり、別の起源であると考えられている。