 ミトコンドリア
ミトコンドリア G2.
グラム陰性陽性細菌・好気性嫌気性細菌について
GIO:
グラム陽性菌と陰性菌の違い/好気性菌と嫌気性菌の違い/大腸菌を好気的環境と嫌気的環境で培養したときのATP合成酵素の役割について調べ理解する。
SBO:
1.グラム陽性菌と陰性菌の違いについて説明できる。
2.グラム染色法について説明できる。
3.解糖系の
ATPの合成について説明できる。4.クエン酸回路の
ATP合成について説明できる。5.大腸菌を好気的環境と嫌気的環境で培養したときの違いを説明できる。
6.化学浸透機構とは何か説明できる。
7.細菌とミトコンドリアの関連性について説明できる。
8.菌の組成について説明できる。
9.好気性菌と嫌気性菌の違いを説明できる。
小山耕太、大串昭彦、北村慶、国重千佳、
浦田雅子、江藤允信、江口太一、大座紀子、植川顕
ミトコンドリアと細菌の関連性
98017小山耕太ミトコンドリア
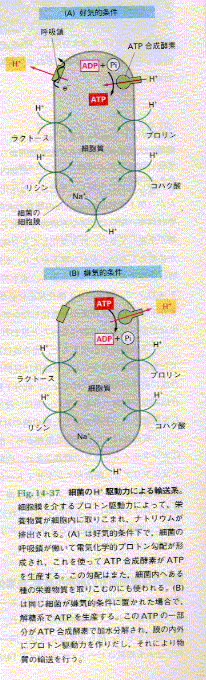
細胞内に存在するカプセル状の小器官。
細胞が活動するためのエネルギーを供給するために、糖や脂肪(→ 脂質)などをアデノシン三リン酸(ATP)というエネルギー源にかえる働きをしている。そこで、ミトコンドリアは細胞の「発電所」といわれる。ミトコンドリアが糖からATPをつくる過程で、酸化還元反応が頻繁におこなわれるため、この働きは細胞レベルの呼吸とよばれる。ミトコンドリアは真核細胞(核が膜の中にある細胞)に存在する。細胞の中にふくまれるミトコンドリアの数は、その細胞の働きによってことなる。たとえば、筋肉細胞のように、とくにエネルギーが大量に必要な細胞は、ほかの細胞よりも多くのミトコンドリアをふくんでいる。ミトコンドリアは酸素を必要とする好気性菌とひじょうによく似ていることから、真核細胞に好気性菌が感染して共生するうちに誕生したと、科学者たちは考えている。
ミトコンドリアの構造
ミトコンドリアは
2重の膜につつまれ、長さは0.5~1μm(マイクロメートル:100万分の1m)、の滑らかな外側の膜は内側の膜と液体でへだてられている。内側の膜は内部にたたみこまれて、クリスタとよばれる構造体をつくり、液体のマトリックス(基質)をつつんでいる。このマトリックスの中にはたくさんの生物学的触媒である酵素や、ミトコンドリアデオキシリボ核酸(mtDNA)がある。mtDNAには直接タンパクを合成するための遺伝情報がはいっている。→ DNA:RNAミトコンドリアの働き
ミトコンドリアのおもな働きは、好気的呼吸をおこなって細胞が活動するためのエネルギーをつくることである。この好気的呼吸では、まず、細胞質内で糖や脂肪が分解されてピルビン酸になり、できたピルビン酸がミトコンドリアにはいって、一連の反応がおこる。その一部がクエン酸回路またはクレブス回路とよばれるものである。
この一連の反応でピルビン酸は酸化され、二酸化炭素と水素原子ができる。できた水素原子は特別な運搬分子によって、内側の膜のクリスタまではこばれる。そして水素は電子伝達系(呼吸鎖)とよばれる膜に結合した1組の酵素・補酵素群にわたされる。
電子伝達系は10個の水素原子をそれぞれ電子(e-)とプロトン(H+)にわける。10個の電子は電子伝達系をすすみ、最後は酸素とプロトンとが結合して水になる。
電子伝達系で電子をやりとりして酸素とプロトンから水ができるとき、大量のエネルギーが放出される。このエネルギーを電子伝達系の5番目の構成成分がつかまえる。この5番目の構成成分がATPase(アデノシントリホスファターゼ)とよばれる、ATPを合成する酵素である。このATPaseは、アデノシン二リン酸(ADP)にリン酸基をくっつけてATPを産生する。このADPからATPができる経路をリン酸化という。
細菌
Bacteria原核細胞(→
細胞)からなる微小な単細胞生物。ふつうは細胞の分裂で増殖する。バクテリアともよばれる。細菌の大半は1~10μm(マイクロメートル:100万分の1m)足らずである。エネルギーの産生や栄養を摂取する方法は多岐におよぶ。空中、地中、水中、氷から熱い温泉まで、ほとんどすべての環境に生息する。深海底の熱水噴出孔さえ、酸素の代わりに硫黄を代謝する細菌のすみかである(→ 海洋生物)。ほとんどすべての食品にみつかるタイプもあり、大半の動植物や他の生物とさまざまな形の共生をする。また、細菌は自分で蛋白質合成もする。だから、ミトコンドリアは一種の細菌であるといえよう。「参考文献」: 百科事典エンカルタ99
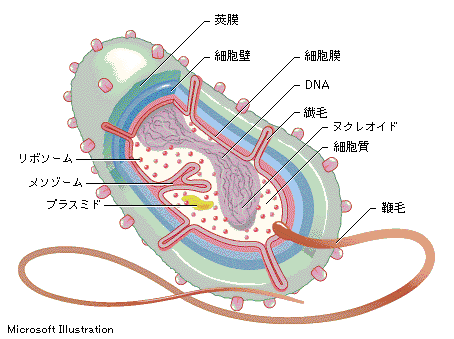
化学浸透機構とは?
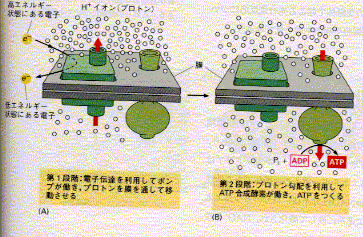
98016大串昭彦細胞内で化学エネルギーの流通をつかさどるのはATPである。真核細胞のATPは、ごく少量が、細胞質中の解糖系により生産され、大部分はミトコンドリア(植物、および藻類の細胞では葉緑体も)の膜に埋め込まれた系によって作られる。細菌の細胞膜でもよく似た反応が行われている。ATPを作り出す機構の基本は生命の歴史のごく初期に出現し、その巧妙な働きゆえに、その骨組みが初期の真核細胞から現存細胞にいたる長い進化の中で維持されてきたのである。反応は2段階からなり、ともに膜に埋め込まれたタンパク複合体によって行われる。
第1段階。食物分子や別の物質の参加で生じてきた電子が、膜にある電子伝達系によって運搬されていく。この電子伝達の過程でエネルギーが放出され、それによって膜の一方向にプロトン(H+)が輸送されプロトンの電気化学的勾配が形成される。このプロトンは細胞中どこにでもある水から得られるが、膜を挟んだイオンの勾配は、エネルギーの貯蔵形態の一つであり、電気化学的勾配に従ってイオンが膜を通過してもとに戻る際にそのエネルギーを仕事に利用できる。
第2段階。H+がATP合成酵素と呼ばれるタンパク複合体をとおって電気化学勾配に従う方向に移動する。ADPと無機リン酸からエネルギーを使って、ATPを合成する反応を触媒するこの酵素は広く存在し、ちょうどタービンのように働いて、プロトンの勾配をを使いATP合成を起こさせる。
この化学浸透機構には、電子伝達、プロトンポンプ、およびATP合成が連携して働いている。
プロトンくみ出しに使うエネルギーを供給する電子は、実にさまざまな材料から取り出されており、好気呼吸でATPを作るミトコンドリアや好気性細菌では、グルコースあるいは脂肪酸の酸化で得られる。化学浸透共役反応では、分子状酸素が最後の電子受容体となり、副産物としては水が生じる。光合成における化学浸透共役では、緑色色素のクロロフィルに光が当たることで必要な電子が得られるし、ある種の細菌では水素や鉄、硫黄などの無機物が高エネルギーを得る材料として用いられている。
化学浸透共役は最初で細菌で出現した。好気的な真核細胞は、その細菌の化学浸透機構をそっくり取り込んだように思われる。おそらく初めは好気性細菌の取り込みによるミトコンドリアの生成、それから藻類や植物へと進化した系統におけるシアノバクテリアの取り込みによる葉緑体の生成があったのだろう。
編性嫌気性細菌も含めてほとんどの細菌は、細胞膜を介したプロトン駆動力を維持している。電気化学的勾配は、細菌が泳ぐときに使う鞭毛の回転運動やプロトンが駆動する
Na
+-H+アンチポートによるNa+の排出に使われる。この系は細菌内へのアミノ酸や糖類など栄養物質の能動輸送にも利用されている。特異的シンポートにより、代謝物質が1個から数個のH+とともに細胞内に運び込まれるのである。細菌の中には、きわめて強いアルカリ性のなかで生存する極端なものもあるが、それでも細胞質のpHを生理的な値に維持しなければならない。こうした細胞では、電気化学的なH+勾配を形成しようにも、逆向きに形成されているH+の大きな濃度勾配に邪魔されてしまう。(細胞内のH+濃度が外部より高い)ある種の細菌では化学浸透に関与するイオンがH+でなくNa+になっているのは、おそらくこのためだろう。このような細菌の呼吸鎖はNa+を細胞外に排出し、輸送系と鞭毛運動はNa+の細胞内への流入によって駆動され、ATP合成も、Na+の駆動するATP合成酵素によっている。このような細胞が存在することから、化学浸透のほうが、普通に見られるプロトン駆動力よりも基本的なメカニズム
 であることがわかる。
であることがわかる。
細菌について
98019 北村 慶
細菌の膜構造と代謝方式による細菌の分類
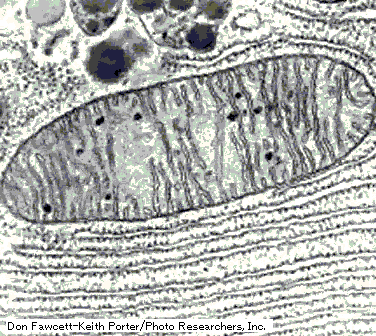
細菌のような原核生物では、原形質(protoplasm)には核や区画化された小胞などは無く、唯一の単位膜系である細胞膜(plasma-membrane)に重要な機能が組み込まれています。
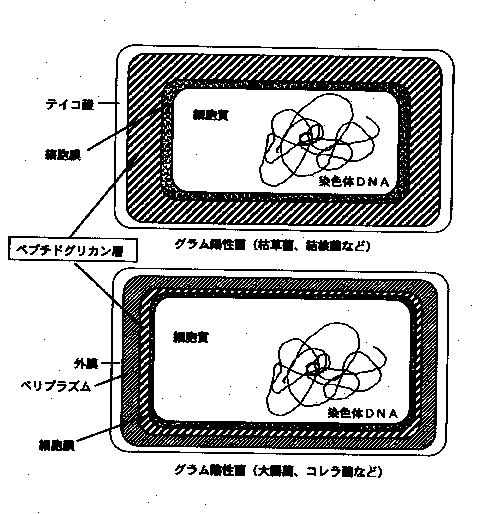 細胞膜は厚さ約 8 nm でリン脂質2分子層から出来ています。リン脂質は2層が逆向きに合わさった形で共に親水基を外側に向けて配列し単位膜を形成していますが、細菌の細胞膜には多様な機能を果たすべく様々な蛋白質が埋め込まれ、こうした蛋白質が膜の乾燥重量の半分以上を占めると言われる程複雑な構造をしています。
細胞膜は厚さ約 8 nm でリン脂質2分子層から出来ています。リン脂質は2層が逆向きに合わさった形で共に親水基を外側に向けて配列し単位膜を形成していますが、細菌の細胞膜には多様な機能を果たすべく様々な蛋白質が埋め込まれ、こうした蛋白質が膜の乾燥重量の半分以上を占めると言われる程複雑な構造をしています。
細胞膜の外側には細胞壁があります。細胞壁はペプチドグリカン(peptidoglycan)という特殊なポリマーから成る網目様の袋で、細胞原形質の浸透圧により細胞が膨張破壊してしまわないように、強靱な構造材として膜を支え、細胞の外形を決定する役割を果たしています。
細胞膜質とは、細胞壁と細胞質を隔てているもので、リン脂質、蛋白質、多糖類で構成されています。
細胞質とは、細胞膜質によって包まれている部分でさまざまな酵素、補酵素、代謝産物、ミネラルなどのほかに蛋白質合成にかかわる成分も含まれています。
核様体は、高等生物の核とは異なり核膜をもたず、DNAの繊維として存在しています。
グラム陰性菌と呼ばれる細菌群では、上記のペプチドグリカン層が薄く、その代わり更にその外側に外膜と呼ばれるもう一つの細胞膜類似膜を有しています。外膜もある種の物質に対する透過障壁となっている為、両膜に挟まれたペリプラズム(
periplasm)と呼ばれる領域は、細胞原形質と外界との中間的な環境になっているものと考えられます。細菌の分類には、デンマークの医師
C.Gram 提唱したバイオレット・ヨード染料による染色性の有無が使われることがあります。クリスタルバイオレットでよく染まりエタノールで脱色されない細菌を 「グラム陽性」、脱色される細菌を 「グラム陰性」 と呼び、これによって原核細胞生物にとって重要な細胞壁(ペプチドグリカン)の厚さを区別しています。 (グラム陰性菌で細胞表層が染料を保持出来ないのは、細胞壁が薄いからという説明と、外膜がある為という2つの説明があります。)代表的な例としては、大腸菌がグラム陰性菌、乳酸菌などの放線菌はグラム陽性菌です。
尚、メタン生成菌や高度好塩菌など一部にペプチドグリカンの無い細菌があり、これらのものは
「古細菌」 (Archaea) と呼ばれ、同じ原核生物ながら分類上は他の細菌(真性細菌 : eubacteria)とは根本的に区別されます。
上記のグラム陽性か陰性かという区別とは直接関係無く、一般に細菌はその代謝面の違いに着目した分類がなされます。
代謝の第一の目的は、エネルギー源となる基質(水素化合物)を酸化して、エネルギー
( ATP ) と還元力 ( NADH ) を獲得することです。この二つを獲得すれば次にはこれを利用して生合成の過程が可能になります。即ちエネルギーを投入しながら同化源基質を還元し、様々な生体高分子を合成し繁殖していくことが出来るようになります。生体のエネルギーは
ATP により供給され、蓄積され、運搬されます。従って細菌がエネルギーを獲得する過程とは、 ATP を合成する、或いはエネルギーを放出した ADP から ATP を再生する過程ということになりますが、その手段には大別して、「光合成」、「発酵」、「呼吸」 と呼ばれる三つの様式があります。(大腸菌が呼吸と発酵を行うように、細菌の中には複数の生育方式を備えている例は珍しいことではありません。)
微生物
(細菌) は栄養源として利用できる基質の別やエネルギーの獲得形態に様々な種類があり、それぞれが細菌の特徴を表すことになりますので、下記のような分類用語によって呼び分けています。
栄養源(細胞合成の為の元素の摂取源)やエネルギー源を、無機物のみに依存して生育出来る自律型の栄養様式を持つ細菌は
「独立栄養細菌」 と呼ばれ、他の生物が作った有機物しか利用できない細菌は 「従属栄養細菌」 と呼ばれます。
生育や増殖の為必要なエネルギーや還元力の獲得は、水素
(化合物) 《水素供与体》 を酸化して行いますが、このやり方について光エネルギーを利用する 「光合成」 方式と、化学反応のみに頼る 「化学合成」 方式の二通りがあります。更に
「化学合成」 方式には、酸素(遊離酸素,化合物中の結合酸素)を酸化剤として利用する「呼吸」方式と、酸素を利用しない「発酵」方式の二通りがあります。「従属栄養生物」( heterotroph )は(例外を除く殆どのものが) 「化学合成」 方式を採り、 「光合成」 方式を採るものは(例外を除く殆どのものが) 「独立栄養生物」( autotroph )です。
「独立栄養細菌」は、外界の無機物(水素、水、硫化水素、アンモニアなど)からエネルギー源となる水素、或いはエネルギー媒体となる一次電子を取る仕組みを持ち、又同時に炭酸ガス
CO2 を取り込んで有機物を合成する代謝系を持っているのが特徴です。(例外的に一酸化炭素 CO を利用して生合成する細菌も存在します。)「独立栄養」
は栄養源として利用する物質の観点から 「無機(酸化)栄養」 とも呼ばれます。一方 「従属栄養」 では原形質(細胞質)に取り込んだ有機物を酸化してエネルギー、還元力、合成素材の全てを得ていますので、 「有機(酸化)栄養」 と呼ばれます。
「化学合成細菌」
が行うエネルギー獲得反応に於いて、エネルギー源を酸化する為に「遊離酸素」を利用する細菌を 「好気性細菌」(obligate aerobes)と呼び、「遊離酸素」を利用しない細菌を 「嫌気性細菌」 と呼びます。
「参考文献」
ホームページアドレス:
http://www.bekkoame.or.jp/i/jackie/microbe/keitai.htmlhttp://www.adguard.co.jp/Discus/science16.html
好気性菌と嫌気性菌
98020国重千佳
|
|
( 「嫌気性細菌」 の内、酸素が毒になるものを 「絶対嫌気性菌」 (obligate anaerobes)、酸素があっても死なないものを 「耐気性嫌気性菌」 と呼ぶ区別もあります。) 好気性菌 酸素の存在する環境でしか生育できない微生物を指す。酸素の存在がその生育に絶対的に必要なものを偏性好気性菌と呼び、カビ、産膜酵母の大部分がこれに属する。乳酸菌などは大気中の酸素分圧より低い分圧の時よく生育し、これを微好気性菌と呼ぶ。 嫌気性菌 遊離の酸素の存在しない環境で生育できる微生物を指す。チトクローム系酵素をもたず、酸素を利用できないのみならず、遊離酸素の存在は有害で、酸素分子20%を含む環境(大気)中では全く発育しないものを偏性嫌気性菌という。酸化的代謝(呼吸)、嫌気的代謝(発酵)を使い分けることができ、遊離酸素があってもなくても生育可能(酸素のある方が生育はよい)なものを通性嫌気性菌といい、細菌や酵母の大部分はこれに入る。嫌気性菌のうち、酸素の存在する環境で生育できる微生物をアエロトレラント・アネローブという。 好気性菌と嫌気性菌の違い 大多数の細菌は、多様ではあるが、ミトコンドリア(や葉緑体)のものと非常によく似たATP合成酵素を細胞膜にもっている。好気性細菌では、電子伝達系がH 参考文献: 微生物学辞典 The Cell
|
グラム陽性菌と陰性菌 98011植川 顕
細菌を光学顕微鏡で観察するとき、色素で染色する。最も一般的な方法が、グラム染色法である。この方法で細菌を染色すると、紫に染まるグラム陽性菌と、赤・桃色に染まるグラム陰性菌に区別できる。この違いは、菌の表層構造上の差異に由来する。
* グラム陽性菌と陰性菌の構造の比較
|
グラム陽性菌 |
グラム陰性菌 |
|
|
外膜 ペリプラズム 膜オリゴ糖 酵素の分泌場所 ペプチドグリカン タイコ酸 多糖体 リポ多糖体 リポタンパク |
なし なし なし 細胞外 多重層 あり 莢膜など なし 一般になし |
あり あり あり(ペリプラズム) ペリプラズム(もしくは細胞外) 単層もしくは薄層 なし 莢膜、スライムなど あり(外膜) 多くの場合あり |
*グラム陽性菌の表層構造
グラム陽性菌の表層は、厚さ約80nmの硬い細胞壁に囲まれている。この細胞壁を構成する物質は、主としてペプチドグリカン、蛋白質およびタイコ酸などである。ペプチドグリカン層は、主として原形質膜を浸透圧のへんかから保護するものである。蛋白質およびタイコ酸の役割はあまり明瞭ではないが、細菌が宿主細胞に接着し、コロニーを形成することに関係があるとされている。
*
グラム陰性菌の表層構造の概略グラム陰性菌細胞膜(原形質)の外側に存在するものは、主として薄いペプチドグリカンと外膜および莢膜などである。陰性菌のペプチドグリカンは、基本的には陽性菌のそれと同じ構造をしている。異なる点は、テトラペプチドのリジンがdiaminopimeric
acidに置換されており、これが別のテトラペプチドのD-アラニンと直接架橋されている。ペプチドグリカンの外側には外膜が存在するが、外膜は陰性菌に特有の構造体である。
解糖系の
ATPの合成 98013江口太一細胞気質で行われる。グルコースがピルビン酸に分解される過程において,2ATPと4水素原子を発生する。嫌気呼吸(酸素を必要としない呼吸)と共通の反応経路である。
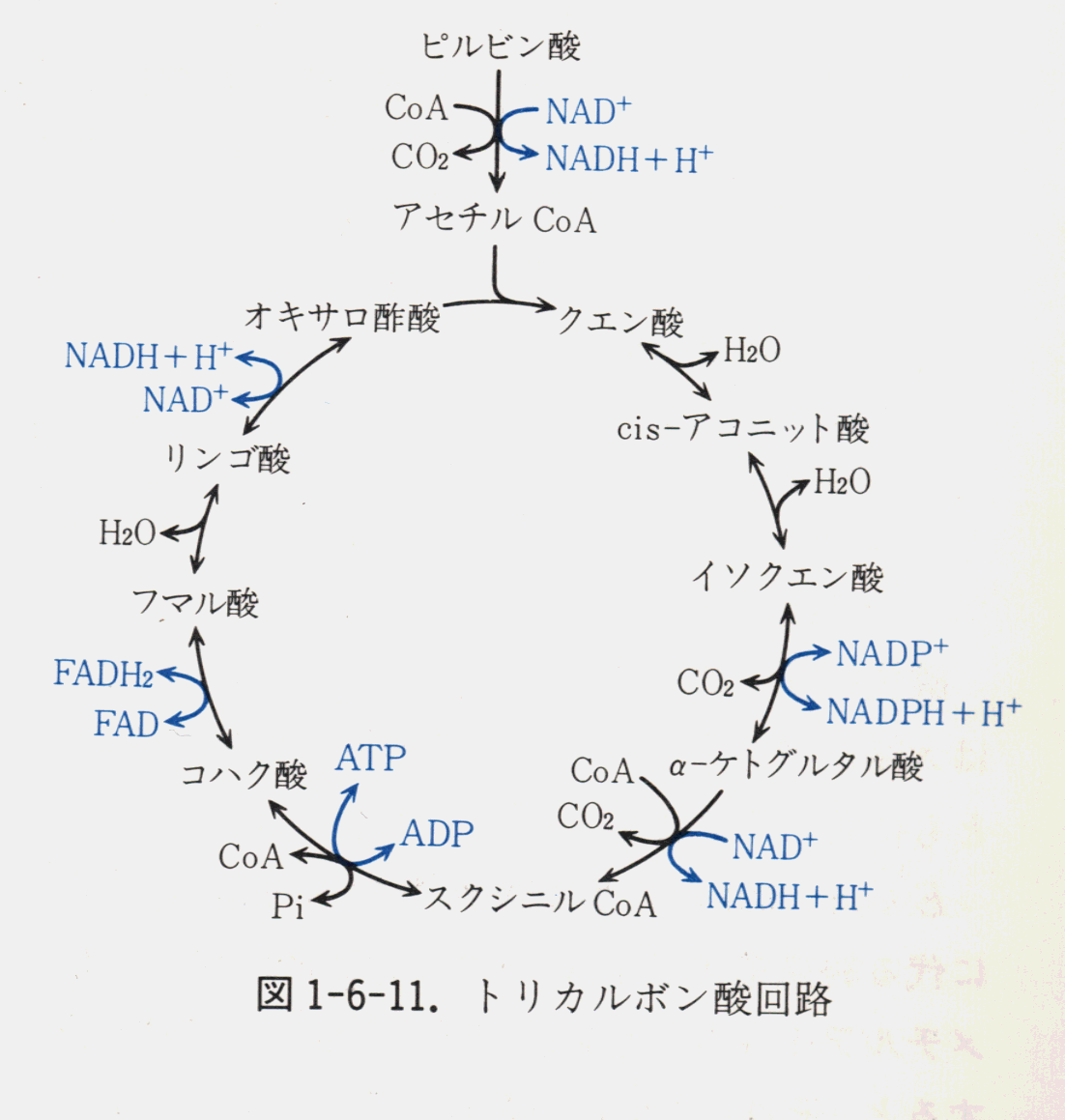
1)TCA回路(クエン酸回路)について
98014江藤允信ミトコンドリアの基質に取り込まれたピルビン酸は、すぐに補酵素A(CoA)と結合してアセチルCoAになる。この過程は単一のステップではなく、触媒する酵素(ピルビン酸脱水素酵素)は3種類の酵素(
pyruvate decarboxylase, lipoamide reductase-transacetylase, dihydrolipoyl dehydrogenase)の複合体である。しかも一つ一つの酵素が複数個あつまった大きな酵素複合体となっている。そのため、効率よくアセチルCoAをつくることができる。 この過程で1分子のピルビン酸から、二酸化炭素が1つ、NADHが1つできる。アセチルCoAは、TCA回路(
tricarboxylic acid cycle, citric acid cycle or Krebs cycle)に入る。TCA回路は名前の示すとおり、複数の酵素反応がサイクルを形成している。簡単にいえば、アセチルCoAはオキザロ酢酸にアセチル基を渡しクエン酸になる。以後、 7ステップの酵素反応で、最初に入ったアセチル基の炭素数である2個の二酸化炭素が放出さ れ、3個のNADH、1個のFADH2、1個のGTPが生成し、ふたたびオキザロ酢酸にもどる。また、2分子のATPが合成される。この酵素反応はすべてミトコンドリアの基質で行われる。 回路に入った2個の炭素は、最初の1回転ではなく、次の回転以降に二酸化炭素となって系 から出て行く。こうして生成されたNADHとFADH2は、ミトコンドリア内膜に埋め込まれた酵素複合体に電子を渡し、この電子は最終的に酸素に渡され、まわりにある水素イオンと結合して水を生成する。このときにNADHとFADH2の持っていた高エネルギーがATPを合成するために使われる。この過程は電子伝達系(electron transfer system)と呼ばれ、28分子のATPが合成される。
98015
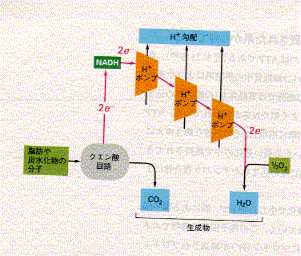
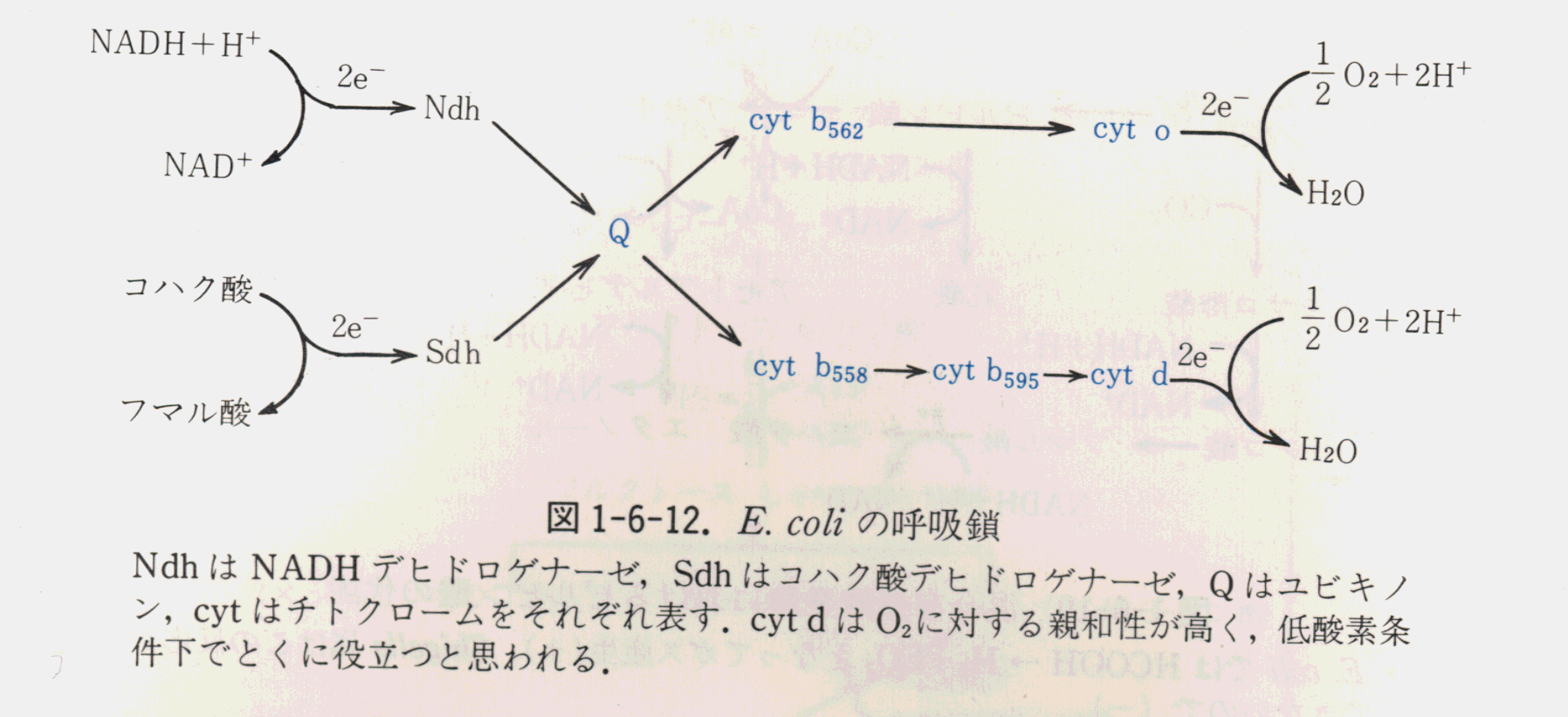
大座紀子大腸菌の好気的環境と嫌気的環境による培養の違い
大腸菌は酸素に対する態度で区分されると通性嫌気性細菌に属している。その中でも、大腸菌は、そのエネルギー捕捉代謝のパターンが酸素の有無に応じて変わらないものである。
ただし、酸素の有無に関係無く増殖が可能とはいえ、エネルギー捕捉代謝の内容は著しい相違を示している。
簡単に言うとO![]() を終末電子受容体とする呼吸鎖が働き、酸化的リン酸化が行われる
を終末電子受容体とする呼吸鎖が働き、酸化的リン酸化が行われる
(酸素呼吸)のであるが、以下の諸点に注意する必要がある。
 還元型補酵素(
還元型補酵素(
好気的増殖の際にも糖類からの酸の産出が見られるのはこのためである。その理由の
一つは、カタボライトリプレッションによって糖類が
TCA回路の酵素や呼吸鎖のチトクロームの生合成を部分的に抑制するためである。
酸化的リン酸化の関与によって
大腸菌は嫌気条件下であっても、
Oこの時、嫌気呼吸を可能にする電子受容体がなければ、菌は糖を混合有機酸発酵でぶんかいして
ATPを調達することになる。