
7班
べん毛の構造と機能と走化性について
G7班
(98060~98068)98060
原田慶美 98065 福田直子98061
東 史子 98066 藤井 可98062
久留有夕美 98067 藤井宏行98063
平野悌志 98068船越亜希子98064
樋渡 敦
GIO
:細菌の持つべん毛の構造と機能およびその生物学的意義を理解する。
SBO
:①べん毛を持つ主な生物を挙げることができる。
②原核生物の構造・機能・役割について説明することができる
③真核生物の構造・機能・役割について説明することができる
④べん毛の化学走化性について説明することができる。
⑤べん毛の化学走化性がべん毛を持つ生物にどのように役立って
いるか説明することができる。
1.鞭毛をもつ主な生物


Dinophyta; 渦鞭毛植物




Euglenophyta; ミドリムシ植物




Euglenophyta; ミドリムシ植物 / Cryptophyta ; クリプト植物



Chrysophyta; 黄金色植物 / Volvocida ; ボルボックス類
= Flagellata
有毛根足虫門(肉質鞭毛虫門)
Sarcomastigophora Honigberg & Balamuth, 1963LINKS
Subdivisions
|
オパリナ Opalinids (Opalinata / Opalinatea / Opalinida) Cepedea, Opalina, Protoopalina, Zelleriella |
種 |
||
|
動物性鞭毛虫 Zoomastigophora |
襟鞭毛虫目(エリヒゲムシ) Choanoflagellida |
150 種 |
|
| ケルコモナス目 Cercomonadida |
種 |
||
| プロテロモナス目 Proteromonadida |
種 |
||
| ハラヒゲムシ目 Retortamonadida |
種 |
||
|
ピルソニンファ Pyrsonymphids (=オキシモナス目 Oxymonadida) |
種 |
||
| マスチゲラ目 Rhizomastigida |
種 |
||
| エブリア目 Ebriida |
種 |
||
| ビコソエス目 Bicosoecida |
40 種 |
||
|
Basal Eukaryotes ディプロモナス目(ヒゲハラムシ) Diplomonadida Giardia lamblia (ランブル鞭毛虫) |
種 |
||
|
Basal Eukaryotes キネトプラスト目(マクムシ) Kinetoplastida Trypanosoma (ネズミマクムシ) |
600 種 |
||
|
Basal Eukaryotes パラベイサル Parabasalids |
ホネマクムシ目 Trichomonadida |
種 |
|
| ケカムリ目 Hypermastigida |
種 |
||
|
植物性鞭毛虫 Phytomastigophorea |
Basal Eukaryotes ミドリムシ (=ミドリムシ植物) |
800 種 |
|
|
アルベオラータ 渦鞭毛虫(オビムシ;=渦鞭毛植物) ヤコウチュウ |
4000 種 |
||
|
クロミスタ カゲヒゲムシ (=クリプト植物) |
100 種 |
||
|
クロミスタ プリムネシウム (=ハプト植物) |
種 |
||
|
クロミスタ: 不等毛植物 ヒカリモ (=黄金色植物;Chrysomonad) |
1000種 |
||
|
クロミスタ: 不等毛植物 ラフィドモナス (=ラフィド藻) Chattonella |
種 |
||
|
緑藻植物 ピラミモナス (=プラシノ藻) |
種 |
||
|
緑藻植物 ボルボックス
|
種 |
||
この他、遊走細胞を形成する菌類、鞭毛菌類がある。このような菌類は鞭毛菌亜門としてまとめられ、遊走子が後端に1本の鞭毛をもつツボカビ綱Chytridiomy-cetes、遊走子が先端に羽型の鞭毛を1本もつサカゲツボカビ綱Hyphocytridiomycetes、
遊走子が側方に2本の鞭毛を持ち、前鞭毛は羽型、後鞭毛はむち型である卵菌綱
Oomycetesの3つがある。(http://www.biol.tsukuba.ac.jp/~inouye/ino/st/mis/colorless_st.html)
References
(福田)
原核生物(細菌)のべん毛について
構造
原核生物の鞭毛は、フラジェリンという蛋白質からなり、真核細胞においての9+2
構造を持たず、機動力は
鞭毛基部の細胞に埋め込まれた部分にある。ここが、モーターとなっている。モーターには、菌体長より数倍も長い、螺旋型をした、鞭毛繊維がスクリューとしてつながっており、受動的に回転することによって推進力を生み出す。
機能
螺旋状に配置した円筒状構造を持つことが知られている。
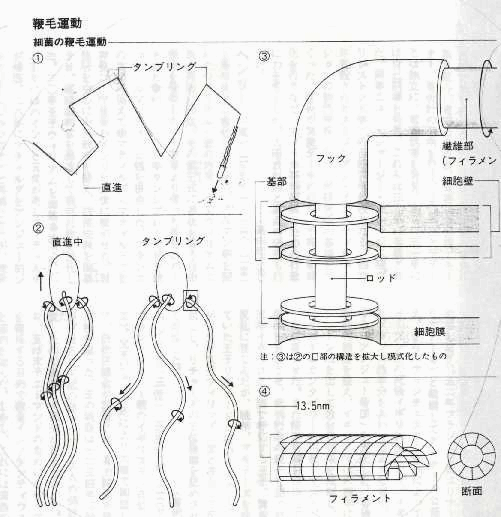
[参考]詳しくはこちら http://bunshi3.bio.nagoya-u.ac.jp/lab/4research1.html http://flux.kz.tsukuba.ac.jp/flagella.html
(久留・平野)
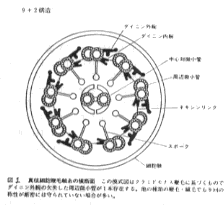
真核生物の鞭毛について
構造
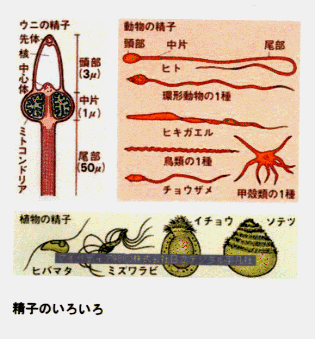
真核生物の鞭毛の基本的な構成は9+2構造と呼ばれているものである。9+2構造とは、周辺二連微小管という小さな管が周囲に9対輪状に、中心単微小管というものが中央に2本(1対)並んだ配置の構造である。精子の場合には種類によってきわめて変異に富み、基本的な9+2構造のものから、その外側に細胞質微小管を持つもの、哺乳類精子のように9本の太い周辺束繊維をもつもの、あるいは中心対の位置に1本の太い繊維をもつもの、6+0や3+0のものなどがある。また両生類の精子のように波動膜を有するものもみられる。鞭毛はふつう細胞1個当り1本か2本で、長さは繊毛より長く数十μmから場合によっては数mmにも達する。直径は0.2μmほどであるが、周辺束繊維をもつ哺乳類精子などではもっと太い。鞭毛の基部は細胞表面下の基粒体に接続している。
機能
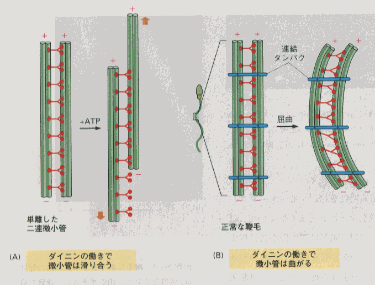 真核細胞の鞭毛運動は、正弦曲線状の屈曲の波が根元から先端へと次々に伝わっていく。繊毛や鞭毛の動きは、微小管が滑り合って折れ曲がることによって生じる。微小管には長軸方向に突き出た多数の蛋白質がくっついており、その中には微小管同士を固定するものや鞭毛を曲げる力を発生させるものがある。屈曲運動を起こすのは、モーター蛋白ファミリーの一つであるダイニン(ATP依存性)である。ダイニンは、尾部をある微小管にくっつけ、頭部は隣の微小管に接するので、二つの微小管に力を生み出す。隣同士の微小管を固定したものが連なっているので、平行に滑るだけの固定されていない微小管の動きと違って屈曲運動が生まれる。
真核細胞の鞭毛運動は、正弦曲線状の屈曲の波が根元から先端へと次々に伝わっていく。繊毛や鞭毛の動きは、微小管が滑り合って折れ曲がることによって生じる。微小管には長軸方向に突き出た多数の蛋白質がくっついており、その中には微小管同士を固定するものや鞭毛を曲げる力を発生させるものがある。屈曲運動を起こすのは、モーター蛋白ファミリーの一つであるダイニン(ATP依存性)である。ダイニンは、尾部をある微小管にくっつけ、頭部は隣の微小管に接するので、二つの微小管に力を生み出す。隣同士の微小管を固定したものが連なっているので、平行に滑るだけの固定されていない微小管の動きと違って屈曲運動が生まれる。
(船越・藤井宏)
4.鞭毛の走化性(
chemotaxis)について
細胞が周囲にある特定の化学物質に、向かっていったり離れたりする性質。
細胞が感知する化学物質の濃度勾配によって方向の定まる運動。
走化性は、単細胞生物の細胞外シグナルに対する応答のひとつである。多細胞動物の細胞間で見られる化学シグナルの伝達機構は、走化性などの単細胞生物が化学的変化に対応する機構を進化させたものが多い。
・真核生物と細菌における化学物質濃度の感知の方法の違い
真核生物では、誘因物質の濃度勾配を直接感知する。
細菌は、時間による濃度変化によって濃度勾配を感知している。
鞭毛の走化性、ということで、以下に細菌の鞭毛による走化性について詳しく述べる。
運動性の細菌は、糖やアミノ酸、小分子ペブチドなどの栄養物質(栄養物質)の濃度の高い方向に向かって泳いだり、各種の有害な化学物質(忌避物質)から逃れる方向に泳いだりする。この比較的単純だがたくみな性質については、大腸菌とサルモネラ菌で詳しく研究されている。

サルモネラ菌はセリンを含むガラス毛細管には誘引され(A)、フェノールを含む毛細管からは逃避する(B)。写真は、細菌を入れた培養皿に毛細管を入れてから5分後に撮影したものである。
細菌は、らせん状の鞭毛を高速で回転させて、運動している。この細菌表面の鞭毛にはらせんの巻き方に向きがあるため、鞭毛の回転の方向によって違った動きになる。反時計回りに回転すると鞭毛は束状に集まり、細菌は一方向へなめらかに動く。しかし、時計回りに回転すると鞭毛はばらけて、方向性のない動きしかできなくなる。
鞭毛は数秒ごとに回転方向を逆転しており、その結果細菌は直線上になめらかに泳いでは突然向きを変えるという特徴のある泳ぎ方をしている。この突然向きを変える動きを「タンブリング」という。
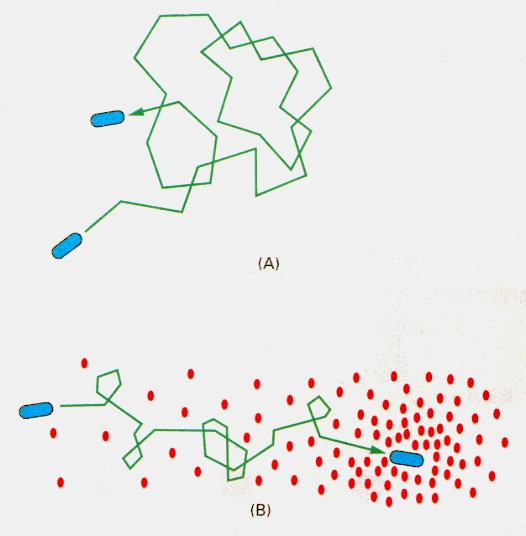
上の図は大腸菌の遊跡であるが、(A)のように走化性シグナルがない場合、なめらかな泳ぎをしてはちょっとタンブリングして勝手に方向転換するので、両者が交互に起こり、三次元的にランダムな動きになる。(B)のように走化性誘因物質のあるときは、その濃度の濃い方へと泳ぐ間はタンブリング頻度が下がり、次第に誘因物質に向かって集まる。こうして、ランダムに動きはするが、指向性が出てくる。忌避物質から遠ざかる走化性も基本的に機構は同じで、逆方向に働くだけである。
先に述べたように、細菌は一定の早さで泳ぎながらある時間内での誘因物質や忌避物質の濃度差を比較することで、空間的な濃度勾配を感じ取っている。実験室で、培地にたとえば誘因物質を加えると、タンブリング頻度は予想どおり数十分の一秒という短時間で減少するが、しばらくすると誘因物質があっても正常値に戻る。細菌は誘因物質の濃度に新たな変化がない限り、この適応状態にとどまる。この適応のおかげで細菌は誘因物質濃度の絶対値でなく変化に応答することができ、誘因物質の非常に広範な濃度での変化に応答できる。
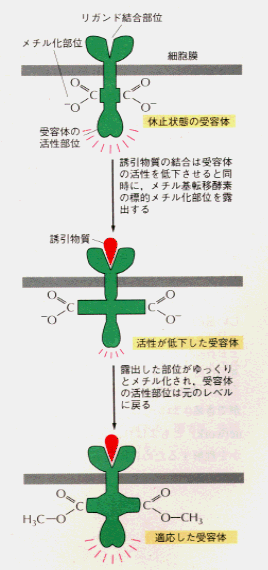
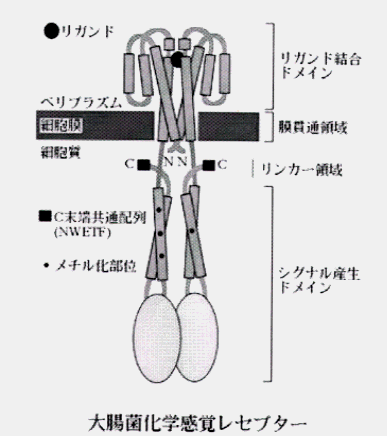
・細菌の走化性の分子機構
いろいろな化学物質に対する走化性は小数の近縁な膜貫通型受容体が膜越しにシグナル伝達を行うことによって生じる。この走化性受容体は適応反応の際メチル化されるので、メチル基結合走化性タンパク(
methyl-accepting chemotaxis protein,MCP)ともいう。受容体の活性は忌避物質濃度の上昇によって促進され、誘因物質濃度の上昇によって抑制される。一個の受容体が、反対の効果を持つ2種類の分子の両方から影響を受けるのである。膜貫通型走化性受容体には4種類あり、それぞれ一群の化学物質に対する応答に関与している。
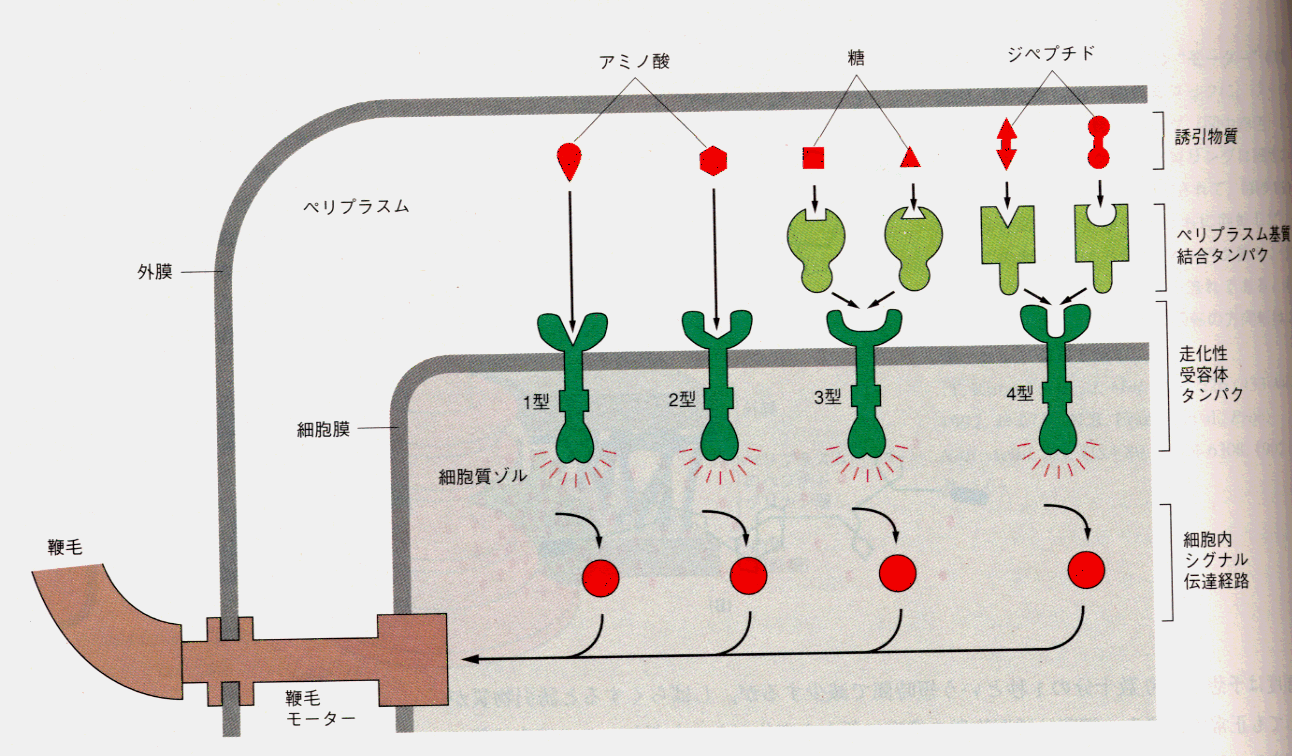
1型:セリンに直接結合
2型:アスパラギン酸に直接結合
3型:ペリプラスム基質結合タンパクを介して糖と結合
4型:ペリプラスム基質結合タンパクを介してジペプチドと結合
誘因物質が走化性受容体に結合すると受容体の活性を低下させ、細胞内のシグナル伝達連鎖反応を遮断し、鞭毛モーターを連続的に反時計回りに回転させ続ける。その結果、タンブリングは抑えられ、なめらかな泳ぎが続く。ココには図示していないが、誘因物質は外膜中の大きなチャネルを通って、細胞外からペリプラスム内へ拡散している。
細菌の走化性で見られる適応現象は、走化性受容タンパクの共有結合的メチル化によって起こる。
(東・原田)
5.べん毛の化学走化性がべん毛を持つ生物にどのように役立っているか
端的に言うと、自分のいる環境を感知し、生育に好ましい場所(糖やアミノ酸などの栄養物に富んだ好ましい環境)や目的の場所に移動し(=誘因物質に近づいていく)、住み心地の悪い場所(老廃物や毒物が含まれた好ましくない環境)からは遠ざかることができる(=忌避物質から遠ざかる)という利点がある。
ここではそれの具体的な例を挙げる。
上記の通り、生きやすいところへ移動する…ということが主なのだが、もう少し詳しく述べてみることにする。
★
大腸菌の場合べん毛を持った細菌の代表として、大腸菌の走化性について見てみる。
大腸菌は4~5本のべん毛を有しており、これが反時計回りに回転するとべん毛は束を形成して菌は直進遊泳し、逆に時計回りに回転するとこの束は形成されずに独立に回転するので菌はタンブリング(tumbling)と呼ばれる遊泳をして方向転換をする。
これを促しているのが、誘因物質・忌避物質などと称される化学物質である。
Adler
らは、数多くの刺激物質に対して、独立に検出可能な化学感覚器(chemosensor)が存在することを証明した。誘因物質に対応する化学感覚器としては、フルクトース、ガラクトース、マンノース、マルトース、マニトール、リボース、トレハロース、ソルビトール、などの糖類、アスパラギン酸、セリンなどのアミノ酸に対応するものがある。
忌避物質に対応するものとしては、脂肪酸、アルコール、疎水性アミノ酸、インドール、低または高pH、硫化鉄などに対応するものがある。 糖のレセプターは細胞外層膜と細胞膜にはさまれたヘリプラスムにあり、マルトース、リボース、ガラクトースのそれぞれに対応するMBP、RBP、GBPと呼ばれるものが知られている。ガラクトースによる誘因は必ずしも糖が細胞内に取り込まれる必要はなく、リセプターに結合するだけで、走化性を生じる情報となり得る。
参考
http://www.ajinomoto.co.jp/ajinomoto/lib/roubun1.htm
大腸菌である彼らが、何を思い、何を求め、何を嫌っているのか、正確に知る術はないが、彼らも彼らなりにより良い生(quality of life)を求めて生きているのは確かであるようである。
精子
有性生殖における雄の生殖細胞。多くはべん毛を持った運動性の細胞で、頭部、中片部、尾部の3部分が区別される。頭部には受精の成立に関係のある先体(リソソームが特殊化したもの)と濃縮された核がある。中片部は中心粒、多数のミトコンドリアを含み、尾部の推進的運動のためのエネルギー供給に働く。精巣で精母細胞の減数分裂によって形成され、DNA、核蛋白質、アルギニンリン酸などが主成分。哺乳類では卵の周囲にあるセメント様物質を溶かすヒアルロニダーゼを含む。卵と合体して発生を開始させ、また雄の固体からの遺伝情報を子に伝える役割を果たす。植物ではコケ植物、シダ植物、イチョウ、ソテツなどが精子を生ずる。
→
精子のべん毛の化学走化性は、我々の生命の起源にも深く影響を及ぼしている、身近なものであるといえよう。精子べん毛運動
→ 生物が受精効率を高めるために獲得してきたシステムの中で、 受精に先立った精子運動の開始・活性化、あるいは卵への走行性といった運動性の変化は、周囲の環境に直接さらされる遊離細胞である精子にとって最もダイナミックな過程の一つである。この運動性の変化は、細胞膜を介したイオンあるいはリガンドの受容とそれに引き続く蛋白質リン酸化/脱リン酸化を主体とした細胞内情報伝達系によって最終的にモーター蛋白質代人やそれに関連する蛋白質が活性化することによって起こると考えられる。
参考
http://www.biology.tohoku.ac.jp/asamushi/inaba-j.html http://www.fujipek.co.jp/~goto/HP/uramain/seishi.htm
マイペディア
98 株式会社日立デジタル平凡社
精子に、卵に引き寄せられる走化性があるが故に、あなたも今、ここに存在していられるのだ。

(藤井可・樋渡)