僿儖儁僗僂僀儖僗偺憹怋
嘆
媧拝偍傛傃怤擖乮fusion丟梈崌傑偨偼endocytosis偵傛傞乯嘇
扙崚乮uncoating乯嘊嘋嘍
僂僀儖僗DNA偺揮幨偍傛傃僂僀儖僗摿堎僞儞僷僋偺抜奒揑崌惉偲偦偺挷愡乮cascade regulation乯
嘐
廻庡僞儞僷僋偺崌惉慾奞嘑
僂僀儖僗DNA偺暋惢嘒
僂僀儖僗棻巕偺宍惉乮nucleocapsid乯嘓
僂僀儖僗棻巕偺惉弉乮nucleocapsid亄envelope乯嘔
曻弌僂僀儖僗偑嵶朎偵媧拝偟丄嵶朎枌偲偺梈崌傑偨偼
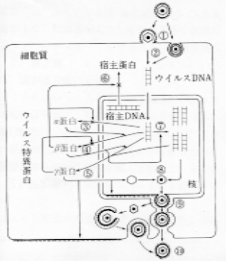
endocytosis乮viropexis乯偵傛偭偰嵶朎幙撪偵怤擖丒扙崚偟偨屻丄 妀撪偱憹怋偟丄妀枌傪偐傇偭偰惉弉偡傞丅僰僋儗僆僇僾僔僪偑嵶朎幙撪偺彫朎枌傪偐傇偭偰惉弉偡傞夁掱傕偁傞乮俠俵倁丄俫俫倁亅俇乯丅
攟梴嵶朎偱偼丄俆乣俇帪娫偺僄僋儕僾僗婜乮埫崟婜乯偺屻丄姶愼僂僀儖僗偼懳悢揑偵憹壛偟丄
14乣16帪娫偱僺乕僋偵払偡傞丅丂
侾丏媧拝丒怤擖丄扙崚
俫俽倁乮扨弮僿儖儁僗僂僀儖僗乯偼僄儞儀儘乕僾忋偺倗俠偱嵶朎昞柺偺僾儘僥僆僌儕僇儞乮傊僷儔儞棸巁乯偵媧拝偟丄倗俛丄倗俢偍傛傃倗俫偵傛傝僄儞儀儘乕僾偼嵶朎枌偲梈崌偟丄僇僾僔僪偼嵶朎撪傊怤擖偡傞丅妀梈崌偵傛傝嵶朎幙撪偵怤擖偟偨僰僋儗僆僇僾僔僪偼妀枌岴嬤朤偵堏摦偟丄扙崚偑婲偙傝丄妀枌岴傪捠偠偰僂僀儖僗俢俶俙偑妀撪偵曻弌偝傟傞丅
丂
俀丏僂僀儖僗俢俶俙偺揮幨偍傛傃僂僀儖僗摿堎僞儞僷僋偺崌惉丒挷愡
僂僀儖僗俢俶俙偼丄嵶朎俼俶俙億儕儊儔乕僛傪棙梡偟偰丄揮幨偝傟傞丅僂僀儖僗摿堎僞儞僷僋偼丄兛乗兝乗兞偺弴偵崌惉偝傟傞乮
cascade regulation乯丅偙傟傜偺僞儞僷僋偼峈尨惈偑偁傝丄偦傟偧傟immediate-early antigen乮慜弶婜峈尨乯丄early antigen乮弶婜峈尨乯偍傛傃late antigen乮屻婜峈尨乯偲傛偽傟傞丅丂
兛堚揱巕偼
5屄抦傜傟偰偍傝丄偦偺揮幨偼丄僞儞僷僋幙崌惉慾奞嵻懚嵼壓偱傕婲偙傞丅兛僞儞僷僋偼俀乣係帪娫偱僺乕僋偵払偡傞丅HSV價儕僆儞乮姶愼惈傪傕偮姰慡側棻巕乯僥僌儊儞僩乮僄儞儀儘乕僾偲僰僋儗僆僇僾僔僪偺娫偺僞儞僷僋乯拞偺兛TIV乮VP16乯偼兛堚揱巕偵僩儔儞僗偵嶌梡偟丄兛堚揱巕偺揮幨傪懀恑偡傞丅兛僞儞僷僋偼偄偢傟傕挷愡婡擻傪帩偪丄兝堚揱巕傪妶惈壔丄兝僞儞僷僋偺崌惉傪懀恑偡傞丅丂
兝堚揱巕偺揮幨偼丄兛僞儞僷僋偺崌惉偑愭峴偡傞昁梫偑偁傝丄俢俶俙崌惉慾奞嵻懚嵼壓偱傕婲偙傞丅兝僞儞僷僋偼姶愼屻俆乣俈帪娫偱僺乕僋偵払偟丄庡偵妀巁戙幱偍傛傃僂僀儖僗俢俶俙偺暋惢偵娭梌偡傞丅兝堚揱巕偵僐乕僪偝傟傞僞儞僷僋偼丄嘆俢俶俙億儕儊儔乕僛側偳俢俶俙暋惢偵昁恵側傕偺
丄嘇僠儈僕儞僉僫乕僛乮俿俲乯丄儕儃僰僋儗僆僠僪丒儕僟僋僞乕僛側偳俢俶俙崌惉偵娭梌偡傞偑嵶朎偺峺慺偱傕戙懼偱偒傞傕偺偑偁傞丅嘆傪寚偔曄堎懱偼憹怋偱偒側偄偑丄嘇傪寚偔曄堎懱偼憹怋拞偺廻庡嵶朎偱偼憹怋偱偒傞丅丂
兞堚揱巕偼丄價儕僆儞傪峔惉偡傞兞僞儞僷僋傪僐乕僪偟偰偍傝丄偦偺敪尰偵偼兛僞儞僷僋偍傛傃兝僞儞僷僋偺崌惉偲俢俶俙暋惢偑昁梫偱偁傞丅偟偐偟丄兞僞儞僷僋偵偼丄倗俛丄倗俢偍傛傃庡梫僇僾僔僪僞儞僷僋側偳姶愼弶婜偍傛傃屻婜傪捠偟偰崌惉偝傟傞傕偺偑偁傞丅
丂
俁丏僂僀儖僗俢俶俙偺暋惢
僂僀儖僗俢俶俙偼丄俿俲偲俢俶俙億儕儊儔乕僛偺堚揱巕傪傕偭偰偄傞丅僂僀儖僗俢俶俙偼丄姶愼俁帪娫屻傛傝崌惉偑巒傑傝丄俋乣
12帪娫帩懕偡傞丅價儕僆儞撪偺俢俶俙偼慄忬偱偁傞偑丄姶愼嵶朎撪偱偼娐忬偲側傝丄儘乕儕儞僌僒乕僋儖條幃偱暋惢偝傟傞丅丂
係丏廻庡僞儞僷僋偺崌惉慾奞
姶愼偵傛傝丄廻庡嵶朎偺崅暘巕慾奞偼慾巭偝傟傞丅
丂
俆丏僂僀儖僗棻巕偺宍惉丒惉弉丄曻弌
僇僾僔僪傪峔惉偡傞僞儞僷僋偼丄嵶朎幙偱崌惉偝傟丄妀偵堏摦偟丄妀撪偱僇僾僔僪傪宍惉偡傞丅儘乕儕儞僌僒乕僋儖條幃偱暋惢偟偨俢俶俙偼僎僲儉扨埵偵愗抐偝傟丄僇僾僔僪偵僷僢働乕僕偝傟偰僰僋儗僆僇僾僔僪偑宍惉偝傟傞丅妀撪偺僰僋儗僆僇僾僔僪偼妀枌傪旐傝堦搙妀枌奜峯偵弌偨屻丄嵞傃嵶朎幙偵怤擖丄僥僌儊儞僩傪妉摼偟丄僂僀儖僗摐僞儞僷僋偱偱廋忺偝傟偨嵶朎枌傪嵟廔揑偵僄儞儀儘乕僾偲偟偰旐傝丄嬻朎撪偵弌夎丄嵶朎奜傊曻弌偝傟傞丅
丂
俇丏廻庡嵶朎偺曄壔
姶愼嵶朎偵偼妀枌晻擖懱偺宍惉偲枌偺曄壔偑傒傜傟傞丅
姶愼弶婜偺晻擖懱偼俢俶俙偵晉傒乮俫俤愼怓偱惵怓乯丄戝偒偔乮廻庡嵶朎偺僋儘儅僠儞傪妀廃曈晹偵埑攔丄妀撪戝晹暘傪愯嫆乯丄僂僀儖僗峈尨傪娷傓乮寀岝峈懱愼怓梲惈乯丅
姶愼屻婜偺晻擖懱偼俢俶俙傪幐偄乮俫俤愼怓偱僺儞僋乯丄妀曈墢晹偺廻庡僋儘儅僠儞偲偺娫偵
halo傪惗偠丄僂僀儖僗峈尨傪娷傑側偄乮寀岝峈懱愼怓堿惈乯丅丂
丂
丂
丂
丂
丂
丂
丂
丂