人体内のマラリア原虫は久しく赤血球内だけで無性生殖が行われるとされていたが、
Short&Granham(1948)らにより組織内の発育(組織型tissue form)が知られるようになり、その後の研究により本原虫の生活史に関する今日の知見が整備された。マラリア原虫は人体内では2つの発育(赤外型、赤内型)からなる無性生殖、蚊の体内では有性生殖が行われる。その生活史について詳しくは別に書くことにする。
スポロゾイト
Sporozoitesの人体内侵入後、肝細胞に原虫が現われるまで一定の時間を要する。肝細胞で多数分裂を行う虫体は無色素性で、三日熱原虫では直径42μmに達し数千個のメロゾイトMerozoitesを形成する。この肝細胞内における発育に要する期間は原虫の種類によるが、およそ1-2週間とされる。この赤外型Exoerythrocytric formにより形成されたメロゾイトはやがて血流にはいり、赤血球に侵入して赤内型Erythrocytric formとなる.赤内型は多数分裂をくりかえし、有色素性である。その分裂により生じたメロゾイトの一部から形成された雌性生殖母体Macrogametocyteおよび雄性生殖母体Microgametocyteが蚊の吸血によりその胃内にはいり、成熱して、それぞれ雌性生殖体Macrogamete、雄性生殖体Microgameteとなる。両者は合体して融合体Zygote、ついで虫様体Ookinete、やがて胃壁の外膜下に出て胞嚢体Oosistとなる。かかる有性生殖により生じたスポロゾイトは唾液腺に集まり、再び蚊の吸血により人体内に注入される。人体内の赤内型は発育分裂(この無性的発育型を一般に繁殖体
Schizontという)を反復するが、その周期は原虫の種類によって一定している。しかも多数分裂にさいして発熱するため、それぞれの原虫種に特有な熱型を示すことになる。
①三日熟マラリア原虫
Plasmodium vivaxもっとも普通のマラリア原虫で、三日熱マラリアの原因となる。世界に広く分布し、日本本土に古くから土着していたものも本種である。
スポロゾイト侵入後、肝細胞内の赤外型の完成に
6-7日を要し、およそ8日後に赤内型が現われる。休眠型(ヒプノゾイト)が存続し、赤内型を補給するため再発しやすく、完全な治療のためには赤外型の殺滅が必要となる。赤内型の発育分裂の周期は48時間で、隔日の熱発作をきたすことになる。もっとも幼若な輪状体
Ring formは直径2.5~3μmで、発育するに従ってアメーバ状運動が盛んになり、染色標本では不正形を呈し、黄褐色の色素額粒がみられる。被寄生赤血球は膨大し、赤紅色のシュフネル斑点Schueffner's spotを有する。やがて核分裂を開始し、赤血球のほとんどを占めるようになった分裂期では通常18-20個の核を有し、その数だけのメロゾイトに分裂する。生殖母体は初めから卵円形を呈し緻密で、色素顆粒に富み、膨大した赤血球の大部分を占める。赤内型増員生殖の周期は48時間である。同期がよいので,患者は48時間毎に発熱のピークを迎える。
②四日熱マラリア原虫
Plasmodium malariae四日熟マラリアの原因となるもので、主として熱帯、亜熱帯地方に限局して分布している。
スポロゾイト侵入後、肝細胞内の赤外型の完成に
14-15日を要する、休眠型は存在しないとされ,再発は流血中に残存する原虫に起因する。赤内型の発育分裂の周期は72時間である。赤血球の輪状体は三日熱原虫よりやや小さく、その後のアメーバ運動も活発でなく、体は一般に緻密で色索に富み、しばしば赤血球上に帯状体
Band formとなってみられる。被寄生赤血球は膨大せず、むしろ縮小することがある。分裂期の核の数は6-12個で、菊花状に並んでいる。生薙母体は三日熱原虫に似ているが、それより小さい。赤内型増員生殖の周期は72時間である。かなり同期がよいので,患者は72時間毎に発熱のピークを迎える。
③熟帯熱マラリア原虫
Plasmodium falciparum主として熱帯、亜熱帯地方に分布し、悪性な熱帯熱マラリアを原菌する。
スポロゾイト侵入後、肝細胞内の赤外型(図ⅠⅠ・55)の完成に
5-6日を要する。休眠型は存在しないとされ、再発は流血中に残存する原虫に起因する。赤内型の発育分裂の周期は36-48時間で、熱型は不定になることが多い。幼若な虫体は他種に比べて小さく、直径約
1.5μmの繊細な輪状体となる。赤血球内に重複感染していることが多い。それが成長して直径約4μmに達したものを大輪状体という。被寄生赤血球は膨大せず、強カに染色するとマウレル斑点Maurer's dotsがみられる。本種では重症感染の場合などを除き、大輪状体以後の発育は内臓の毛細管内で営まれるため、末梢血管には認められない。生殖母体はバナナ状または半月状で、半月体Crescentといわれ、その完成したものが末梢血液内に現われる。本種には,薬剤耐性がもっとも古くから認められている(というか,最近,三日熱マラリア原虫にも耐性種の存在が確認されるまでは,唯一の薬剤耐性をもつ種だった)。早くも1910年に,ブラジルでキニーネ耐性が報告されている(Peters, 1982)。現在では,クロロキン耐性,アモディアキン耐性,ファンシダール耐性の他に,メフロキン耐性やハロファントリン耐性のものも報告されている。赤内型増員生殖の周期は,個々の原虫については48時間であるが同期が悪いので,患者の発熱は明確な周期性をもたない。
④卵型マラリア原虫
Plasmodium ovaleまれにみられるもので、主として熱帯アフリカに分布し、卵型マラリアを原因する。
肝細胞内の赤外型の完成に約
9日を要し、三日熱と同様に休眠型が存在するとされる。赤内型の発育分裂の周期は48時間で、三日熱と同様の熱型を示す。赤内型の特徴は四日熱原虫に似ているが、帯状体はみられない。分裂期の核の数は
6-12個である。被寄生赤血球はやや膨大し、しばしば卵型で、一端が鋸歯状を呈することがある。またシュフネル斑点類似のものが早くから見られる。生殖母体も四日熱マラリア原虫に似ているが、赤血球が少しく膨大した斑点が著しい。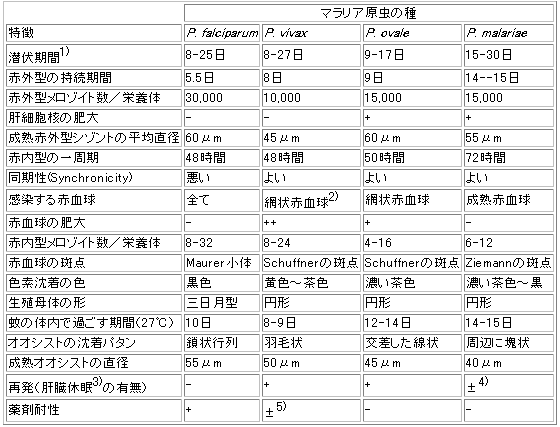
1)
感染力のある蚊が刺してから原虫の出現(塗沫標本で検出されるようになる)までの期間。
2) Reticulocyte
。赤血球の生成過程としては,まず骨髄で正赤芽球(normoblast)が盛んに分裂し,次第に細胞体が小さくなり,血色素を合成するようになり,ついで核が萎縮して細胞分裂を止め,RNAが減少するとともに血色素合成が増加する。その後脱核が起こって扁平になったものが,網状赤血球である。網状物質は細胞質にわずかに残っている細網状のRNAで,brillant cresyl青で染色される。この細網がなくなったものが,成熟した正常赤血球である。血流中の赤血球のうち,0.3~1%は網状赤血球で,赤血球造血の盛んなときはこの割合が増加する。
3) Hypnozoite
といって,肝臓ですぐには分裂せず休眠状態で存在する原虫があらわれる。三日熱マラリアと卵形マラリアで再発が起こる原因は,かつては肝細胞内で反復される第二次赤外型の存在が想定されたが,1978年から1982年にかけてのKrotoskiを中心とする米英の研究者の集中的な研究の結果,それは完全に否定された。それにかわって,\strut 蚊から注入されるスポロゾイトの中にただちに肝内で増殖を開始するものと,一ヶ月から数ヶ月を経てから何らかの原因で増殖を開始する「肝内静止型」の二種類あることが証明された。この「肝内静止型」の存在は,理論上はShuteによって1946年に予言されていたが,実証的にはKrotoskiが蛍光抗体法を用いて,1979年にアカゲザルでサルマラリアの一種であるP. cynomolgiについて,ついでP. vivaxを感染させたチンパンジーで示し,後に組織培養のヒト肝臓癌細胞でも示され,確実なものとなった。
4)
肝臓のヒプノゾイトはないが,赤内型が30年も持続することがあるので,とくに治療が不完全な場合に再発はおこりうる。例えば,脾臓摘出手術の36年後に再発した報告がある(Tsuchida et al., 1982)。
5)
最近,薬剤耐性のものが報告された。例えば,パプアニューギニア高地の居住者がかかった三日熱マラリアでは,塩基にして2400 mgのクロロキン投与でも改善が見られなかった(Schuurkamp et al., 1992)。さらに,パプアニューギニアでは,メフロキン耐性の三日熱マラリア原虫も報告された(Amor and Richards, 1992)。同じくパプアニューギニアでは,プリマキン耐性のヒプノゾイトも報告されている。