免疫
この問題は,マラリアワクチンの潜在的なターゲットである。マラリア原虫の生活環のうち,ヒトの血清中の抗体に曝露するのは,スポロゾイトとメロゾイトだけである。ヒトの体内での他のすべての時期は細胞内で生活し,免疫的防御の発現から守られている。生殖母体が蚊の中腸内で活性化されて,生殖体が放出されると,蚊が吸い込んだ血液中の抗体に曝露するけれども,通常特異的抗体は存在しない(生殖母体は,ヒトの血流中では細胞内に存在する)。実験では,生殖体のタンパク質に対する抗体は,蚊への感染をブロックする。
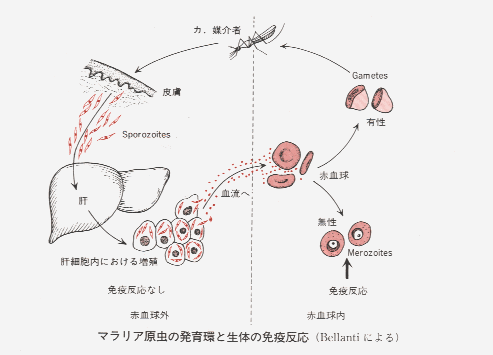
マラリアに対する先天免疫は、たとえば人が鳥のマラリアに感染しないこと、アメリカの黒人が白人よりも三日熱に感染しないなどの事実から、その存在が知られる。
マラリアの免疫は一般に赤内型に対するもので、マラリア原虫の反復感染を受けると、後天免疫を獲得するようになる。かかる獲得免疫は一種の感染免疫で、少数の原虫が体内に存在する間だけ免疫が持続し、治療などによって完全に原虫が消失すると免疫を失うようになる。,抗体産生系を破壊する
HIVウィルス感染者でのマラリアの研究結果から,マラリアが風土病である地域で成人がマラリアを発症しにくいのは,抗体ができるからではなくマラリア原虫への特異的キラーT(TC)細胞による細胞性免疫が増強されるためだと報告された(Butcher, 1992)。マラリアの抗体はγ
-グロブリンのIgGに多く含まれており、各種の血清反応に利用される。高度のγ-グロブリンを有する濃厚マラリア浸淫地の成人血清に治療効果があることも認められている。〈マラリア感染時に細胞性免疫が増強される過程〉
ヒトの免疫は,抗原提示細胞(周辺域マクロファージ,単球/マクロファージ,濾胞樹枝状細胞,樹枝状細胞,ランゲルハンス細胞)が,自己の
MHCのクラスIまたはクラスII抗原と同時にその異物のエピトープ(短いポリペプチド鎖)を膜表面に提示し,それをヘルパーT(TH)細胞が認識することによって特異的に活性化される。 TH細胞は,B細胞を補助して抗体を産生させたり,TC細胞,ナチュラルキラー(NK)細胞,マクロファージ,顆粒球,抗体依存細胞障害(K)細胞など,さまざまなエフェクター細胞の作用を調節している。これらTH細胞の作用の多くは,リンホカイン(インターフェロンなど)によって媒介されている。しかし,他の細胞とくにマクロファージから遊離されるサイトカインも重要な役割を果たしている。また,T細胞とB細胞の働きはともにサプレッサーT(TS)細胞によって調節されている。マラリア患者では感染初期に,血清中のサイトカインの一種である,腫瘍壊死因子
(TNF)の濃度が有意に上昇することが知られている。これは,マクロファージが貪食作用によって原虫をとりこみ,分解するときにTNFを放出するためである。すると今度はTNFがマクロファージを含むエフェクター細胞群に影響を与え,THからのリンホカインなしでもマラリア原虫のメロゾイトを殺すようになる。
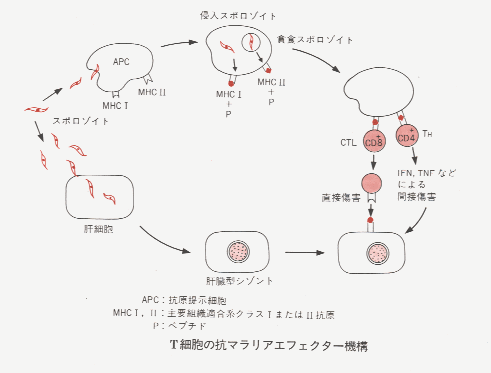
細胞性免疫の存在を示すものとしては、従来からマラリアに感染すると細網内皮細胞が増殖し、脾、肝、骨髄の食細胞が被寄生赤血球、遊離した原虫、色素などを盛んに摂取することが認められていた。また、患者の流出中のリンパ球が熱帯熱原虫に対してある程度発育を阻害する作用を有することなども、その存在を示すものである。